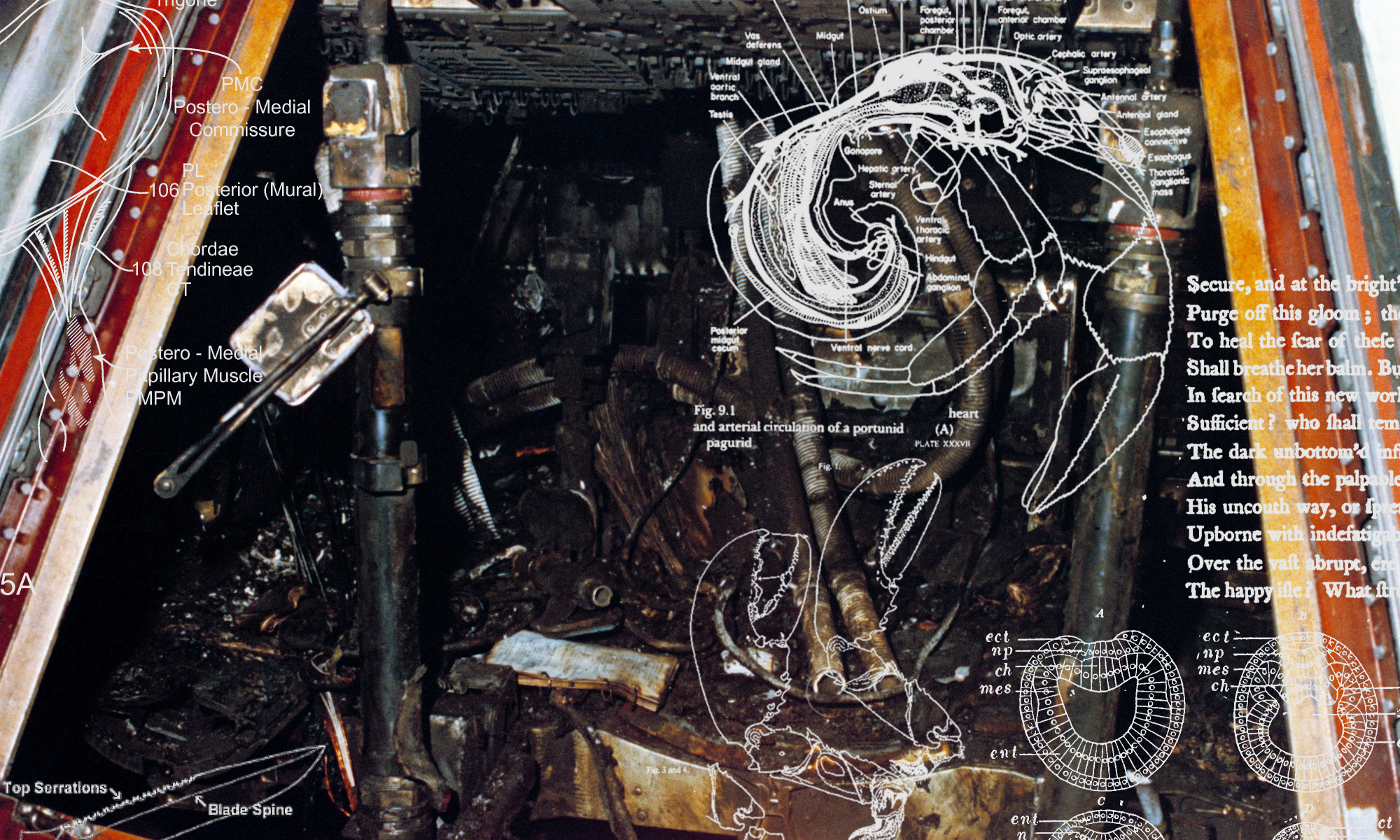

by pps
‘One of their philosophers has lately discovered that “as the liver secretes bile, so does the brain secrete thought”; which astonishing discovery Dr Cabanis, more lately still, in his ‘Rapports du Physique et du Moral de l’Homme’, has pushed into its minutest developments. […] He fairly lays open our moral structure with his dissecting-knives and real metal probes; and exhibits it to the inspection of mankind, by Leeuwenhoeck microscopes and inflation with the anatomical blowpipe. Thought, he is inclined to hold, is still secreted by the brain; but then Poetry and Religion (and it is really worth knowing) are “a product of the smaller intestines!”'[note]Thomas Carlyle, ‘Signs of the Times’, in Edinburgh Times (1829).[/note]
Coeliac Splanchnogenesis, Nervous Speleogenesis, Intellectual Epitheliogenesis: or, the Art of Sinking Inside Oneself
All enfoldings, invaginations and internalisations attendant upon abiogenesis are only so many precursors and ancestors to the later development of intelligence’s full-blown transcendental functioning. However: this is not to say they are therefore the same. It is not to speciously state their dubious ‘continuity’ — whether explanatory or descriptive, genetic or categorial. They resonate only in mutual dissonance. For the ‘transcendental’ is not an ontological-descriptive feature (despite being gregariously read as such in various strands of philosophy), but an essentially normative-functional one. To apprehend it properly: in becoming responsible to what is called transcendental, one folds oneself into a nexus of accountability that is accountable, in a special way, only to itself. Thereby, it is, importantly, irreducible. It is a collapse inwards, into itself, that cannot be reversed or reduced to anything else. But, in the sense that it is a collapse into itself (the very model of irreducibility and saltation), it echoes the lining of a gut, the deposition of an epithelium, the nervous recurving of encephalisation. Invaginating into a linguistic nexus echoes the inward folding of a coelom, not by therapeutic continuity but by reciprocal saltationality, because they are both perfect enclosures. They proceed by creating potentiating blockages against the external world.[note]Our purpose here is not to flatten thought into some kind of continuity with non-thinking processes and thus arrive at therapeutic immanence (via a narrative of dubious inheritance or recapitulatory reverie). Thinking and matter (whether the matter be organic or not) are discontinuous. (Thinking was never ‘contained’ in previous nature, and it never will be — indeed, its essential nature is to strive against this, to ramify and inflame this discontinuity — and it did not evolve or unfurl from the latent possibilities of some ‘vibrant matter’ or ‘vitality’.) Indeed, our purpose is thus merely to point out that it is the very nature of the thing we call ‘evolutionary’ development to create irreversible discontinuities: its nature is to be discontinuous with itself. By not being itself, nature triggers life; by not being itself, life triggers thought. Life is reality’s attempt to exit itself, this is why it exists only via exits, it is a saltus generator.[/note] And so, clinging to regulative argument as in-built deniability or alibi against flagrant abuse of analogy, let us embark on a phylogenetic fantasy.[note]We follow the visionary model of the speculative master, Sigmund Freud. Freud, A Phylogenetic Fantasy: Overview of the Transference Neuroses (HUP, 1987).[/note]
Novalis, a long time ago, proposed the historico-philosophical study of a “DIETETICS OF MANKIND”.[note]Novalis, Notes for a Romantic Encylopaedia (SUNY, 2007), 14.[/note] So too Nietzsche once exclaimed: “Verily, my brothers, the soul is a stomach!”[note]Nietzsche, Thus Spake Zarathustra (CUP, 2006), §16.[/note]
One imagines the transcendental architectonic as a metabolic system: Sensibility providing catabolism of uncategorised exteriority into manageable chunks through sensory mastication; Imagination as the filtration that synthesises materials into a digestible manifold; Understanding as the anabolic process that builds matter up into Intellectual Nourishment;[note]John Milton wrote that the Apple was “intellectual food”: thus, we must eat it to the core. Cf. Sellars, ‘The Structure of Knowledge’ (1975), http://www.ditext.com/sellars/sk.html[/note] the Table of Categories, perhaps, as Table of Anabolic Enzymes.
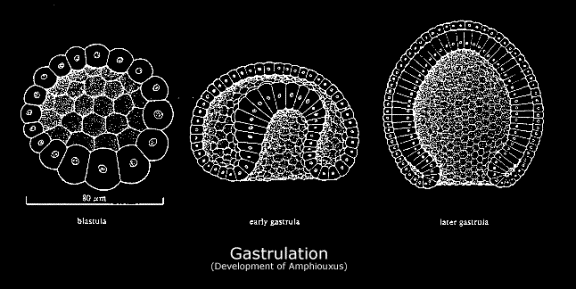
Before we had central nervous systems, we had colons. This applies at both ontogenetic and phylogenetic levels (in embryogenesis, the blastula folds itself into a gastrula[note]Gastrulation is the complexification of a blastula into a trilaminar embryo consisting of the three layers of the ectoderm (outside/distal layer), mesoderm (middle layer), and endoderm (internal/proximal layer). (It is an internal differentiation, or schistosity, wherein the successive embryonic laminae come to echo the geognostic-stratigraphic structure of the planet, or, the foliation of strata as noticed by those like Nicholas Steno or Gottlieb Werner.) The ectoderm develops into epithelium or skin, the mesoderm weaves itself into muscle and bone, the endoderm transforms into the digestive system and viscera. Notably, it is during this phase that the embryo folds into itself to create the alimentary canal: sculpting the mouth and anus and the uninterrupted tract between the two. These are formed first via the proctodeum and stomadeum, which are depressions that invaginate into the anus and mouth, respectively. The digestive system is then created via endodermic evagination: a kind of internal hollowing out (splanchnogenesis echoes speleogenesis echoes intellogenesis). During this stage, the coelom (or primary body cavity, designed to house the viscera) opens up within the embryo: an internally directed speleogenesis, or opening up of a cave, this is carved out via a process known as enterocoely. Note that the cover image to this post depicts a gastrulating zebra fish embryo.[/note] before it neurulates into a recognisable chordate). Arising far further back in the phyletic archaeology[note]Again, not stressing continuity, but transition and saltation.[/note] of what has now developed into language-using and reason-wielding navigation, digestion is merely the first form of ‘locomoting’ an external world via the installation of a productive blockage or boundary against it. Just like the ‘proto-transcendental’ thrown up later by the CNS sensorium (and we strive to be delicate with our analogies here),[note]A CNS is ‘proto-transcendental’ because it is a representing of the world, in the sense of a contrivance or manufacturing, thus creating a stark separation within the world between appearance and reality; however, it is not a fully transcendental structure because this requires the even stronger separation between justifications and causes. This only emerges with the arrival of language, via concepts and thus normative structure. The transcendental is full-blown normative: a CNS only produces differential dispositions, it cannot justify their reliability.[/note] the digestive system is a potentiating blockage that allows for selective navigation of external modalities (a selective uptake of the outside world that potentiates an ability for discriminative locomotion).[note]Mobility taken here in an abstract sense of the ‘locomoting of various modalities‘ is not necessarily spatial (for example, the selection and uptake of nourishing materials as opposed to non-nourishing can be taken as a form of navigation of environmental modalities). Abstractly, then, it refers to the locomotion of a possibility space or space of options. Spatial locomotion, indeed, came subsequent to the locomotion of chemical co-ordinates and gradients instigated by metabolic economy. Most basically, locomotion means anything that is an election or optimization for certain modalities above others, rather than mere passivity. ‘Locomotion’, in this sense, may provide a suitably minimal descriptor for ‘life’.[/note] It therefore intensifies the individual qua individual, allowing for a separation from the world that ramifies navigation of it.[note]More precisely, by creating a productive block (by largely managing to block out much of the external stimuli — much like the damming of a river), a principle of discriminative uptake is generated (damning creates utility). Now, because there is a distinction between ‘inside’ and ‘out’, there can be discrimination of what gets in. Finitude — material or cognitive, energy economy or attentional economy — is the prerequisite of orientation, for one can only navigate the world when one comes to successfully de-laminate, or separate, oneself from it: the ability to discriminate can only arise within the generation of constraints that necessitate the need for discrimination. As Kant put it: “I orient myself only according to a subjective ground of differentiation”. – Immanuel Kant, ‘What Does It Mean to Orient Oneself in Thinking?’, in Religion and Rational Theology, ed. & trans. Wood & Giovanni (Cambridge: CUP, 1996), 9.[/note] This is ‘locomotion’ in an abstract sense: as the optimization for certain modalities, be these the navigation of gastrochemical co-ordinates within the culinary universe rendered by the gut, or of individuated objects within the spatiotemporal universe only later rendered upon the arrival of the CNS. In both cases the potentiation of individuality is only bequeathed via a productive blockage — a separation — that is thus revealed as the early ancestor of the later transcendental: not because of explanatory continuity but, we again stress, because of a reciprocal structure of saltation via inward collapse.
Noogenesis is preceded by splanchnogenesis as the first finitude-generating intensification of ‘internality’ that, whilst further entrenching the formative disjunction between organic-system and externality, also thereby galvanizes the organism’s empowerment over this external landscape. Paradoxically, it is only through exiting the world (by further collapsing into itself: twisting and torquing into itself to form a gut in endodermic evagination, or retreating into the simulative encasement of a nervous system) that the organism comes to grasp this world as a world. The invagination of an alimentary canal or, later, the bilaterian centralization of nervous nets into a CNS can bring a ‘world’ into view — an organismic umwelt — only through limiting, or blocking out, external stimuli.[note]Worlds need bounds to give them structure. This is a basic Kantian insight: that it is the basic conceptual (rule-governed) structure of cognition that bequeaths this (without a cogniser, there is no world). Kantian discursive-judgement moulds the cascade of Humean sensation into a whole. ‘World’ is a necessary, i.e. functional and regulative, feature of inquiry. [/note]
The evolution of bilateral anatomical symmetry (itself the sufficient condition and precursor to a centralized and segmented nervous system) is another example of this: by dispensing morphological radiality — as panoptical immanence and immersion — the promotion of only one plane of symmetry[note]Bilaterian morphology drops symmetry on the transverse and coronal planes, only retaining it on the so-called sagittal plane. Through this we give up a circular or spheroid form, and gain an identifiable ‘front’ and ‘back’. Worms and vermicular lifeforms — as the simplest bilateria — are perfect examples of this: they are oriented entirely by a mouth (front) and anus (behind), and thus appear to be merely animated digestive tracts.[/note] generates an orientational ‘front’ and ‘behind’ for the organism, whilst also promoting the intensifying localization of sensory glands into a ‘face’ or ‘front-end’.[note]Mandibles unfurl, teeth regiment, the mouth — portal to the organic universe — opens onto the world. From this bilaterian invention of ‘faciality’ (the evolution of the splanchnocranium), the entire sensorial universe bubbles outwards and backwards from the mouth as a knotting reflexion of nerve-concentration. Eyes and nasal apertures open as world-interfaces bubbling upwards from behind the mandibles, emerging, originally, in order to help guide culinary items down into the esophageal labyrinth. Considering the CNS as a functional protuberance upon gut purposiveness, the major sensory organs unfurl — flower-like — backwards from around the mouth, the splanchnocranium only later bulging back into the neurocranium.[/note] This morphological genesis of faciality thus further lifts the organism out of immersion: by blocking out peripheral fields through the reinforcement of a directional aperture, it foreshadows the arrival of a ‘perspective’ in the world (once more, an empowering separation from the world that facilitates locomotion within it via selective uptake). As a directionalisation of the sensorial field through perspectivally filtrating peripheral foci (localizing the sensorial universe into a directed cone), it creates an invisible structuration of the organism’s universe that is simultaneously a further separation from externality. This is why the development of bilateral symmetry and faciality is likewise another precursor (yet only a precursor, and regulatively speaking) to the ‘conceptual transcendental’, in that it provides conditions of objectivation that cannot themselves be objectivated (unless we use a mirror, or, alternately, the mirror of explicating language): an invisible and necessary structuration of reality that is also simultaneously a filtration, a separation, from it.[note]Kant himself relates our bilateral symmetry to the production of space as form of intuition in his discussion of our handedness, or ‘chirality’ (i.e. the fact that, although our hands are identical in shape, they can never be superimposed one over the other: a right-handed glove can never fit a left hand). Kant uses this to argue that our experience of space cannot be reducible to abstractions or relations. Even further, it implies that our experience of space must be derived from our position in the world (nothing else could decide which hand is ‘left’ and which is ‘right’). In this way, Kant opens up the isthmus through which transcendental psychology touches upon spinal anatomy and, further back, the loss of radial symmetry. We orient ourselves in the world only via a subjective ground of differentiation. However, the physiological ground of this ‘subjectivation’ does not therefore belong to humanity; nor does it belong merely to mammalia or even to chordata; we share one of these initial planes of orientation with echinoderms, with arthropods, with nemotodes.[/note] The evolution of bilateral symmetry created the conditions under which predation could flourish (this truly is “fearful symmetry”, burning bright, in the forests of the night): faces are markers of lethality, of the dispensation of a pre-lapsarian organic radial communism; it symbolizes the increasing relinquishment of life to the telic, temporally-productive structures that progressively come to shape all terrestrial development.[note]Life lost its claims to immanence with the loss of radial symmetry: with bilateral directionality, life came to anatomically resemble the secret teleonomies that come, increasingly, to puppet it. Bilateral symmetry presages the installation of modernity because it feigns the installation of end-orientation and teloi. It is the physical translation of organic tropism into an anatomical plan. Striving, condensed and coagulated into a body-plan. The loss of radial body-plans is the biotic equivalent of the Fall; nevertheless, it gave us eyes with which to see, noses with which to smell, and — most importantly — teeth with which to tear.[/note] By blocking out peripherality (or, radial immersion), an individuated — focused, directed, hungry — perspective on the world is produced; this is a delamination from externality that actually empowers the organism’s locomotion of externality by allowing for the prioritization and de-prioritization of external stimuli (‘your prey is in front of you, ready to be clasped with your forward-facing mandibles’).[note]A representation that represents everything is not a representation, but instead the suffocating and undying unity of Parmenidean autarchic identity. Only through informational limitation or compression (i.e. mediation) does representation occur.[/note]
This limitation is precisely what synthesizes a ‘world’ in the first place (where previously there was only indistinction and blind continuity).[note]As Kant noticed, there was a big difference between ‘world’ (which arrives, always, as categorially pre-structured) and ‘noumena’ (the total lack of categorial structure). Moreover, the enunciation of ‘externality’ presupposes internality: so, without blockage you do not even have this outside.[/note] Locomotion, and later ‘perspective’, can only come with finitude.[note]These blockages thus emerge as the phylogenetic ancestry to our robust phenomenological sense of spatiotemporal ipseity — of nowness, of hereness.[/note] You need an aperture to view the world (just as you need a crucible in order to recreate it), and the first such aperture was pharyngeal: nervous-differential and discursive-normative apertures came later on (later, GUIs). Although both are methods of navigation, digestion retains vast temporal precedence over representational means and, indeed, for the majority of its — relatively brief — history, this latter has been entirely subservient to digestive forms of exchange.
The first ‘world’ was thus gastrochemical, only arriving to us through an oesophagus. It is only very recently that the second, sensory ‘world’ (of objectivated spatiotemporality, siphoned through a sensory nervous system) has managed to lift itself off from its functional substrate: taking off into auto-complexification as a form of inner-ramification (like the inward torquing of a complexifying gut). Thus, all forms of representation (both sentient and, even, sapient) have, for the largest part of their history, been merely functional appendages to food-acquisition. The organism’s CNS-derived ‘global-simulation’ of itself and its environment began simply as an overgrown side-effect of nutritional contestation (because better spatial locomotion allowed for surer procurement of nourishment, initiating a sensory arms-race that was locked-in — upon evolutionary arrival — by the mutually enforcing dynamics of predation and anti-predation counter-measure). Sentience and, much later, sapience begin life as a self-exaggerating excrescence of the stomach.
From this (limited) perspective, the sensory manifold is therefore a later and, originally, functionally subservient information layer merely superimposed on top of our primary, peristaltic thoroughfare. (Indeed, sapient intelligence in humans was likely first emancipated from this functional subservience to splanchnic ends by the invention of cooking which, via externalising digestion and outsourcing gastric labour, freed up surplus energy to be re-invested into a meandering process of roundabout development, otherwise known as ‘civilization’.)[note]Cf. Wrangham, Catching Fire: How Cooking Made Us Human, (Profile Books, 2010).[/note] One notes, through this, that the first way that we, organisms all, interacted with the world was through our guts: eyeballs with which to see came only later; and, then, only as a way to locate our prey; which was itself a development of our evolved ability to eat each other. The first ‘world’ (‘world’ understood as self-propagating and mutually individuating division between inside and out) was a gut; the second ‘world’ — of time, space, and objects — was derived only as a ramification, an originally functionally-subservient information overlay, added on top of the first. Long before we rendered an objective world within the informational englobement of a CNS or the self-correcting procedure of discursive inquiry, we carved out a gastric world for ourselves deep within our primary body cavity, or coelom. And, for the longest time, the latter functionally enveloped the former. The sphere of the sensorium was preceded by the coelom; Plato’s cave was preceded by a splanchnic speleoplex.
Continuing this train of thought, one might be pleased to consider intelligence primarily as functional excrescence: mere protuberance or apophysis of gut-function; one that, turning into itself, became self-catalyzing; and, if one were so inclined, one could include the whole story of human history within this tumorous loop. Sensorial thoroughfares (eyes, nose, ears) bubble upwards and outwards from the mandibles and splanchnocranium (as merely functional appendages to an alimentary tube); subsequent to this, the creation of the CNS’s global simulation balloons backwards into the expanding neurocranium (intelligence swells backwards from the mouth into a skull) as a physiological divagation from primary gut-function (a branching that originally serves the gut, to better represent prey) and, finally, the installation of a symbolic-linguistic encasement arrives as result of this cranial ballooning’s rebounding unto itself, through increasingly localised swelling of prefrontal cortex (providing expansion of working memory, thus facilitating grounds for executive function, advanced goal-direction, and aptitude using linguistic prostheses).[note]Coolidge & Wynn, ‘Working Memory, its Executive Functions, and the Emergence of Modern Thinking’, in Cambridge Archaeological Journal, 15:1, 5-26, (2005) & Ambrose, ‘Coevolution of Composite-Tool Technology, Constructive Memory, and Language: Implications for the Evolution of Modern Human Behaviour’, in Current Anthropology, 51:1, (2010).[/note] (This opening up of working-memory provided the niche for ensuing memetic invasion/symbiosis.) The grand process of encephalisation is an ontological erring, echoing the original Adamite error (again, intellectual nutrition).
The interface chauvinism — unique to us as bilaterally symmetric animals — which presumes that CNS-derived world-interfaces (the electric vagaries attendant upon congeries of overgrown ganglia) are the only ways we locomote the world, forgets this enveloping gastric ur-relation, universally shared by metazoan life. And yet, this is mere pleasing dream — a myth. For chauvinism is our special fate. We are eternally shut off from any such ur-relation and its therapeutic isthmi: for, just as bilaterian predation rescinds claims to radial community, sentience abjures any claim of functional flattening. ‘[F]or after the first bite there is no return to innocence’.[note]Wilfrid Sellars, ibid.[/note]
Like the archaic gut itself, the very adaptive-functional success of sentient overlay (its ability to further nutrition acquisition) lay in its collapse into self-accountability (as potentiating blockage): a sacrifice of immediacy for interminable mediacy, in the informatic invagination of a nervous system, that in collapsing into itself becomes able to represent its states to itself. And, later, in the holistic-hermetic recurving into the ‘inside’ of a linguistic nexus that finds authority and warrant only in itself, such that it can rebuke, if it wish, the passions of the gut. In these ways, these features are propelled intensively beyond their origin, as escape velocity, from original status as exaggerating functional-appendage. Thus, resonance only in dissonance.
For, just as metabolism is only capable of extracting energy by siphoning off externality — admitting it only in homeopathic and reconstituted doses — so too is the CNS only capable of representing legible structure by shutting the organism into its own hermetic simulation or model of the world. (There is no ur-relation but self-relation.) Nervous systems galvanise navigation not by giving us contact with a pre-existent reality, but precisely by shutting it off: enfolding the organism into holistic self-relation, such that it can represent its own states to itself. Perspective is generated through blockage: the filtration of externality through the alimentary tract; the localization of peripheral fields in faciality; the englobing computational constraints of CNS simulation; the irreducible conceptual encasement of glottogony. ‘Immediacy’ is thus replaced, forever, with interminable self-mediation, locking the organism into its own modellings. No turning back.
Accordingly, the nervous system is the result of the informatic ‘invagination’ of a piece of the world into a globally-enclosing simulation of itself and its environs, such that it can only ‘read’ its own stimuli[note]Through this informatic invagination, the system comes to be able to represent its own states to itself (it begins to be able to ‘read’ its own outputs as inputs). External stimuli thus still affect (or ‘feed’) the system, but they can only become legible by first becoming part of it. Thus, unmediated externality — or immediate contact with an externality — is forever shut off by the development of a nervous system. This was the requirement — the entry-fee, let’s say — for a world of appearances to be able to arise. Because the nervous system is the introduction of representation into the world it is also the introduction of mediation, and thus also ancestor to finitude.[/note]: the post-chordate universe arises as each organism shuts itself into the internally-constituting prison of its own representations. Once again, this is why — in carefully regulative terms — the CNS is a veritable ‘ancestral echo’ to the full-blown linguistic-normative transcendental, despite the absolutely unbridgeable gap between the two: because, through informatic invagination, it can now only ‘read’ its own output as input, and thus it comes to pre-empt Kantian finitude and synthetic a priori judgement whereby all knowledge only proceeds through relation to the conditions of knowledge. It also provides a precursor to language as a hermetic system of signs that refers only to itself and its own structures and conventions, rather than anything outside of it. This separation from the world is what lends it its transformative power: it allows for involuting manipulation and self-correction of the system; or, it contains no authority that is beyond criticism, no datum beyond update.
The CNS provides a similar function at the organismic level. Because it represents reality how it seems rather than how it ‘is’ (i.e. through a layer of irrefragable mediation), it can optimize this representation for the organism’s needs and requirements.[note]The ‘is’ of reality would be a computational avalanche and impossible cascade that would be utterly infeasible to internally represent or filtrate: just as the idea of deriving nutrients from everything, from matters of all backgrounds both biogenic and inorganic, is equally preposterous.[/note] Of course, it is crucial to note, that this ‘optimising’ proceeds in irreducibly different ways between the two: for the CNS, it proceeds due to adaptive-selective-pressure; whilst, for language, self-correction proceeds by way of regulating norms of discourse. The latter allows optimisation to take place, moreover, at intensely faster speeds, because it allows the process to selectively intervene within itself (rather than relying on the stochastic processes of natural selection as in CNS simulation, for example). Put simply, both the CNS and language hold no unmediated contact with the outside world, both are a globally self-enclosing re-creation of this world, that therefore contains no ‘transparency’[note]Transparency here is not meant in Metzinger’s unique usage as a form of ‘interface blindness’ by which a model mistakes itself for a reality (cf. Being No One, 2003): rather, it is intended in the more intuitive sense of denoting an unmediated or unmitigated contact with outside reality. I.e. total perspicuity or the lack of interface refraction.[/note] (the veritable birth of simulation).[note]The idea, or phenomenological feeling, that such a system, be it linguistic or nervous, does contain transparency — i.e. contact with some outside world — is the trick that the nervous system plays on itself: reality is the feeling created within a simulation that is not able to self-represent the fact of its own nature. It is a unique form of ‘interface blindness’, wherein the interface is itself entirely invisible to itself. Reality (in the sense of naïve realism, or, immediate contact with externality) is merely the result of such computational limits. Again, cf. Metzinger (2003).[/note] But this blockage is, again, why both are so powerful: both create the selective uptake — the discrimination of information — that structures externality into a world in the first place. Just like the linguistic normative transcendental, the CNS, as ancestral ‘echo’, moulds the formless cascade of external stimuli into an objectivated universe (of, for example, individuated objects) that can therefore be usefully locomoted by the organism (selective uptake allows for optimal intake). It is a not a window onto some pre-existent reality but, rather, more accurately a miniature (and representationally self-sufficient) universe within the universe: a mitosis or budding off of reality — an organismic chronotope. Finitude, then, as ‘both poison and cure’.[note]Wilfred Sellars, Ibid.[/note]
The evolution of digestion, of faciality, and of nervous simulation, therefore constitute the intensification of self-relation: the internality they procure is nothing other than an increase in reflexity. This is why the base-plan for an organism is the sphere: because the sphere — since at least Parmenides’s mobilization of the image — has become the geometrical paradigm and tautegorical symbol of self-reflexivity. Each point on the sphere can be antipodally related to another point, without exception: it is, thus, entirely self-enclosing.[note]Accordingly, transcendental morphology deduced that all life was essentially spherical: the German anatomist, Friedrich Tiedmann, wrote that “[a]ll organic bodies, plants as well as animals, have a form more or less round and oval”, composed of “convex and concave surfaces” (whereas “inorganic bodies” are “limited by flat surfaces and right [angles]”. cf. Friedrich Tiedemann, A Systematic Treatise on Comparative Physiology, Introductory to the Physiology of Man, trans. James Gully & James Hunter Lane (London, 1834), 17.[/note] Note that the braincase is spherical (thereby recapitulating the globe that it stands open and models or maps for itself). The more the organic system comes to relate to itself, the more it intensifies itself as a separation (delamination) from external thoroughfare (self-serving self-relation is individuation). As the distinction between outside and inside comes to feed back into itself as the form of its own self-intensification, separation from the world (as the intensification of separation — the installation of ever more elaborate divorcements from continuity, ever more extreme encasements) gradually becomes self-entrenching.
The organism doesn’t just propel itself within the world, it also — at evolutionary scales — propels itself into itself as self-escape: first, more complex guts, for better extraction of energy; second, a sensorial-nervous encasement to lubricate this extraction; subsequently, the qualitatively unparalleled pitch into a hermetic linguistic-inferential nexus of responsibilities; finally, and premised upon the prior, the enabling enclosure of the organism into a conceptually-saturated universe. Each level of increasingly flamboyant incarceration is at the same time an unprecedented emancipation and potentiation. Poison and cure: self-intrication is self-extrication. (This is the true and proper meaning of Sir Thomas Browne’s resonant intonation: “Thus is man that great and true amphibium, whose nature is disposed to live, not only like other creatures in divers elements, but in divided and distinguished worlds”. It is also translates in the realm of ethics into the conviction that increasing autonomy stacks with increasing responsibility.) And this process proceeds because, with each step and threshold of self-extrication, the increased motility bequeathed by this extrication is skimmed off — as a form of leeway or modal drift with regards to claustrophobic identity with externality and its attendant causal tyranny[note]The ancestor of heteronomy.[/note] — which (as a form of unmooring or delamination) is looped back, or re-invested, into the extricating process in order to propel further escape.
This is, again, ‘motility’ in a domain-agnostic sense, applying at each respective level of modality: first, gastronomic discrimination, then spatio-temporal locomotion, and, finally, in the nimble tracking of entitlements and responsibilities as rational ‘scorekeeping’.) The mobility bequeathed by unmooring is looped back in order to render more unmooring, thus further mobility. The more we explicate, the more we can explicate. This self-directed escape, we stress, is only ever generated as further inwards collapse (progress is quicksand): as progressive self-incarcerations, progressive self-entanglements into increasingly complicated internal universes of representation (the more a perspicacious sensory world appears in front of us the larger the neuronal machinery grows behind-the-scenes; the more our discursive know-how expands the more intricated within an expanding conceptual and normative nexus we become). The organism runs away from reality and itself — ‘swimming upstream’ — by inventing ever more self-intricating, ever more complex prisons… and each time, it throws away the key, making ‘immanence’ an ever more receding fever dream of ‘escape’. (Of course, this evidently makes the lie of immanence ever more attractive.) From this perspective, the organism is not so much a sphere as a labyrinth (one that builds itself).
Like the elastic energy stored within a coil spring, the organism (as iterative separation/process of immanence secession) loops back into itself, accumulating potency after potency. With each gyre, it invests the built-up energy back into itself in order to intensify its involution, coiling infinitely inwards like a nautilus.[note]At some point, however, thermodynamics dictates that all the coils will unwind. All the labyrinths will solve, or escape, themselves.[/note] Lethality builds up behind the face, fangs serrulate back into the throat, deadliness coils into itself as internal complexification, mesodermal differentiation, enetercoely, ossification, consumption, predation. Such self-investment is a model for abiogenesis — the stochastic origination of a boundary that, in turn, creates an energetic gradient between inside and out, a gradient that, subsequently, somehow comes to feed back into itself as a form of self-propulsion. Life is a biotic Ponzi scheme that invests in itself and itself only — a deepening of itself as itself, an unmooring from the external ‘world’, a collapse inwards. This ‘self-investment’ also therefore arises as the prehistoric blueprint of temporality: an intensive difference between present and future, created in the progressive orientation of the organism towards its own intensification as an emergent ‘goal-directedness’. It is the transmigration of life’s self-propelling gradient from a chemical domain to a properly temporal one that makes it identical to the production of time itself.[note]At least, time in its strictly empirical, or more generally CNS-specific, sense.[/note] It does not matter that ‘teloi’ — and, later, ‘reasons’ and ‘responsibilities’ — do not actually exist in externality, because — regardless of this — they become real when part of the world starts to act as if there are such things. A regulative ideal is thus a reality infection. Part of reality comes to decouple itself from identifying with itself as itself — i.e. as claustrophobic uniformity, as dead matter — and opens up the hole through which temporality—as self-directedness through self-alterity — leaks. Unrealities begin to impinge on reality, futures begin to distort the present: history creates itself.
Intelligence as Meontotaxis
The more life separates itself from externality, the more a world — as a globally enveloping structuration of appearances — comes into focus for it: the more life involutes and self-complexifies, the more it internally generates the structures of ‘proto-finitude’ within itself that potentiate a structurated — spatially and temporally undergirded — world. Inward collapse rebounds on itself as the generation of an internally-constituted world (just as nervous centralisation rebounds on itself as cephalisation and the creation of a reality-budding chronotope/simulation). Paradoxically, it is this collapse inwards that enables the ‘outside’ to arise as an ‘outside’.[note]This reaches its zenith in knowing. For, only by separating ourselves from ‘reality’ can we come to know it: this is what representation is. And representation is an unavoidable, irreducible, aspect of knowing. Kant showed this best: knowledge is not just produced in spite of the boundary installed by finitude, it happens because of it.[/note] More optimised digestive systems require further folding inwards and endodermic involution; more eidetic representational sense-worlds require further involution into the CNS (by furthering dependency on an internally-constituting information system). Metabolism utilises an energetic gradient (between inside and out) to potentiate the extraction and storage of work; representation, likewise, feeds on the productive split between appearance and reality (produced by a blockage, generated by the creation of mediation in nervous simulation[note]Mediation consists in a break from identity: wherein something resembles something without being the thing it resembles. Mediation starts when reality begins to reproduce itself via dissimulation (first taking place within the knotting of nervous ganglia). Nevertheless, it is precisely the subsidiary nature of this reproduction — its status as ‘simulacra’ — that grants it its potentiating, or empowering, quality: for, because it is not identical with what it reproduces, the mediated representation is able to provide the unmooring that allows for manipulation and optimisation (i.e. the precipitation of a ‘world’ from the filtration of environmental information from environmental noise). The gap within reality that allows parts of reality to start to delaminate from brute givens and come to select and de-select certain representational episodes. It is within this self-intensifying chasm — between appearances and reality — that intelligence itself is generated (intelligence considered as the ability to become aware of thoughts as thoughts, and thus to optimize them — to unlock the ability for self-optimization or self-intervention — rather than encounter them as incorrigible parts of reality). Again, progression is generated when parts of reality trick themselves into behaving like they are not parts of reality.[/note]) in order to filter externality into relevant ‘worldly’ information.[note]This ‘gradient’ (the split between internal representation and noumenal reality) becomes productive for the organism because it can represent the world insofar as it is relevant to its goals (of nutrition acquisition, reproduction, etc.). It becomes self-productive, moreover, because the complexification of the sensorium (the CNS sense-world) only arrives via increased ‘locking-in’ of the organism into its own universe: further exaggeration of the distinction between ‘appearance’ and ‘reality’, in order to produce ever more complex and ever more eidetic world-models. At cognitive levels, the complexification of our representational world only comes through further traction and entanglement in a transcendental architecture. Theatrical verisimilitude is granted only via the complexification of artifice (the behind-the-scenes machinery exponentially swelling and ramifying in step with increase in verisimilitude). The very same applies to the generation of a world in consciousness — and cranialization, the expansion of our ossified brain-case in order to accommodate the increasing complex neurophysical rendering of this ‘world’, is the physical record and ledger of this. Indeed, this is why every attempt to escape the world (via representation, involution) simultaneously further weighs the organism (as an attempt to escape itself) down within it. The more we create an eidetic second reality, as a simulative attempt at escape via organic self-involution, the more our brains — and their energetic requirements — swell. Simulation has a real-world price. Empowerment over the world — achieved via escape from immanence with it — is always like quicksand. The more life escapes into its own universe, the more it becomes implicated and complicated within the universe beyond its epithelial, nervous, and conceptual encasements. We are just not intuitively aware of this (thus, the illusion of ‘transcendence’) because it was never economic for a brain to simulate itself as a brain (i.e. a glucose-hungry organ, rather than a perspicuous window onto some pre-existent, pre-structurated reality). It was never economic for the simulation to model its physical basis. (But this is to say more than is necessary at this point.)[/note] The self-intensification of these gradients (requiring further divorce from externality as the auto-catalysis of interiority) thereby becomes self-feeding. Separation from the world feeds back into itself as a form of its own self-propulsion (the evolutionary inception of the generation of temporality, but also the glimmerings of autonomy as the ability to track the partition between ‘what is’ and ‘what is not’ in order to align ourselves to the ‘right’ or ‘lawful’): the complexification of the organism’s own internal universe issues from ramifying the organism as an implosion into itself, a process that becomes its own self-installation.
Life, thus, can be seen as reality’s tortured attempt to escape itself. This is why the organism evolves anterior-posterior asymmetry: in order to give it the directionality by which it can run away from itself (and this is why we continue to hate our bodies: misosomatology goes deeper than any genealogy of morals, bubbling up from the bedrock of abiogenesis itself). Yet, like an elastic band, each attempt at self-escape rebounds the animal into itself, as auto-complexification, with redoubling force. The organism, as escape trajectory, is thus a strange form of reality-denial: indeed, in thermodynamic terms, this is indisputable; but, so too, is the concept-monger’s attempt to re-shape the world in terms of sanctions and laws; for the latter is inherently a form of orientation towards non-being (though ‘reality-denial’ and ‘non-being’ are no longer baldly pejorative, here; for, as we have seen, the powers of ‘denial’ or ‘blockage’ are potentiating). The intensification of deficiency is unparalleled empowerment: disentangling itself from determination, disintrication from brute existence simultaneously bequeathes the spaciousness for maneuever that becomes self-looping. As Herder loved to note, a human is a “Mängelwesen”: a creature of deficiencies. The complexification of organic life sets off an attendant race inwards, a self-internalisation, an exit from the world that — paradoxically— also empowers the organism’s worldly performance: the stomach begins this, ossification empowers it with the bony enclosure of viscera, and encephalisation — literally — crowns it with ganglion diadem. However, self-internalisation does not stop here, it merely transports itself — in the dawn of noogenesis and glottogony — into a fully transcendental-conceptual-normative domain. This, however, is what connects digestion to intelligence: they are both iterations of reality’s escape from itself via the deepening of separation from identity. Life identifies with itself as an abrogation of identity: a self-feeding negativity. As a deepening of self-relation, noogenesis emerges as a further entrenching of the original schism initiated by metabolic function. Self-relation is the very paradigm of separation from externality — the progressive delamination, or exit, from external causal structures bequeathed by something turning inwards to become its own primary causation (or, in the case of conceptual cognition, its own sole justification or reason). Life tends towards causal holism and then towards rational hermeticism. It is an attempted exit into justificatory onanism. Thus, rather than being a mere functional excrescence, life’s inwards collapse into intelligence merely represents a continuation of the tendency initiated by metabolism: a continuation, however, only in discontinuity.
This is not to say that digestion and noesis are, in and of themselves, ontologically continuous: this is not an argument for some kind of nutritive monism, it is simply to note that these processes are analogous in that they both intensify the formative discontinuity between organism and world. The irreducible ‘fall’ into inferential hermeticism echoes the irreducible ‘fall’ into nervous mediation, and echoes only in its irreducibility. Precisely by becoming capable of grounding itself only in itself rather than as a means-towards-nutrition, the genesis of reasons represents a ramification of the discontinuity that the evolution of metabolism first itself instantiated and nervous systems extended. The potentiating blockages that began with the development of metabolism eventually blossom forth in the full-blown installation of an insuperable barrier to experience that simultaneously generates the very possibility of said experience. To expand: Hume long ago decreed that legitimate knowledge was not possible beyond immediate sensible intuition; Kant countered that indeed this was possible, but only via a self-relation (the mathematician can make synthetic a priori judgements because they refer only to possible intuitions regarding space or time, in geometry or arithmetic respectively). Thus, it came to be realised that knowledge only is through a self-containment, or self-limitation — a blastulating envelopment.[note]Analytic apodictic statements are not truly knowledge, because they are tautology. Thus, in the synthetic a priori, Kant made knowledge productive in an involuting fashion: it learns by explicating itself more and more, and it drives itself towards this. It continues the trend of collapsing inwards.[/note] Kant showed that, without a conceptual structure to relate to, the mere cascade of intuitions could not generate, or be organised into, ‘experience’; this conceptual enclosure thus creates epistemic content — precisely by limiting the raw materials of intuition by way binding them to what can be inferentially justified about them and what they inferentially justify (thus generating a new, non-somatic form of ‘interiority’ via an inferential holism). Because of this infolding, implexing holism (no epistemically contentful judgement is not justified, or mediated, by further judgements), knowledge (the ability to retroject reasons for epistemic content and project the reasons said content enables) generates itself precisely by folding into itself and closing itself off against the ‘world’ (i.e. there is no self-justifying knowledge generated through unmediated contact between thought and being).[note]In other words, when we become enveloped within concepts, the ability for ‘knowledge’ is finally generated (the soundness of claims can become measured against their conceptual aptness) through and within this separation from the world (knowledge can only be justified by further reference to concepts, creating an inferential holism that cannot be escaped from: in other words, there is no bridging between the world and knowledge).[/note]
Conceptual transcendental entanglement triggers knowledge by creating a sphere of legitimacy through limitation: it is — currently — the crowning productive blockage. Kant was certainly sensitive to this aspect of self-enclosure. He often compared reason to a sphere (just like the ossified englobement of the cranium, the gastric cavity of the coelom, etc.). Indeed, he claimed that, although the “earth as it appears to [one’s] senses” is merely a “flat surface”, we can, in “accordance with principles a priori”, know that it “is a sphere” with “circuit, magnitude, and limits”; and, accordingly, he stressed that exactly the same applies to cognition, because our “reason is likewise not alike” to a “plane indefinitely far extended” but “must rather be compared to a sphere”.[note]Immanuel Kant, Critique of Pure Reason, trans. M. Weigelt (London: Penguin, 2007), 606-8.[/note] This metaphor demonstrates that, although the content of sensible intuition is potentially infinite (in the sense of a sphere’s boundless two-dimensional surface), the space of reasons governing it has englobing limits (just like the spatially finite, three-dimensional sphere). And so, just as the ectodermal deposition of skin is what individuates the organism from the buffetings of externality, we see that the epigenesis of categories and concepts is what provides the enclosure of finitude that marks out the reasoning subject, or, perhaps better, the reasoning community.
Certainly, language-acquisition (the process through which the subject first becomes ensconced and intricated within the architecture of concept-mongering) can be analogically seen as the deposition of a normative exoskeleton (parallel to the prenatal englobements that enclose the organism within its own universes).[note]This ‘exoskeleton’, of course, being unique in the fact that it is collectively constructed and updated by a community of agents.[/note] Moreover, by inventing an entirely novel order of justificatory entailment (of reasons, as opposed to mere causes), the installation of this conceptual-enclosure represents the largest saltation from externality yet furnished. The upshot, basically, is that human organism locks itself off into a hermetic linguistic-conceptual universe. However, just like its precursors, this lock-in arrives as an unparalleled empowerment for, and potentiation of, navigation, precisely because of its retreat from the tyranny and claustrophobia of immanence, the intensified saltation introduced between ‘appearance’ and ‘reality’ (bequeathed by conventional symbols and their arbitrary nature) actually allows for an extended flexibility of actions. By not being fully anchored in reality, the human organism therefore comes to find creative solutions to ancient evolutionary problems. Indeed, neurophysiological evidence of this productive unmooring or delamination is found in the fact that homo sapiens, even when compared to their hominid brethren, present a high degree of ontogenetic neoteny (the retention of paedomorphic — or childlike — traits later into development). Essentially, this means that the human brain arrives — and remains — underdeveloped, thus underdetermined or lacking cemented structure, and therefore exhibits the plasticity required for unmoored (i.e. creative and discontinuously novel) cognition. A creature of deficiencies, umbilically connected to ‘non-being’: that ‘GREAT AMPHIBIUM’. Camped between the domains of spacious non-being and exigent being. Neoteny reiterates the peeling away from ‘that which is’, or mere being, that finally matures into advanced intelligence and its ability to locomote, and even reinvent, the structures of possibility itself.
We can now, taking speculative leaps, see how life — as a self-intensifying escape from reality via inward collapse — is reality’s attempt to escape itself through the progressive unmooring of itself from claustrophobic and suffocating identity with itself. This unmooring is facilitated by the ontological introduction of mediation (through productive barriers) that trigger an ontic liquidation effect: for, as reality increasingly becomes unlike itself (in its revocation of uniformity or identity), discontinuity and change are gestated as the inner turbulence (meontic drift) that is evolutionarily registered as the organism’s increasing extrication of itself from heteronomous causal tyranny. This extrication consists in the process whereby the organism comes to relate primarily to itself, and thus to cause itself, and later to justify itself, eventually sealing itself off into its own spontaneous justificatory-explanatory order. Such causal hermeticism instantiates modal drift: the possibility for new possibilities. We have seen how this, as the birth of self-directedness or end-orientation, could be said to have generated time (or — at the very least — chronoreceptivity: which, from a certain perspective, is — of course — identical to the production of time itself). However, with the birth of the conceptual order, life’s auto-investment in its own intensive gradients emigrated from both energetic and chronogenic dimensions by coming to finally colonise and extend itself into a fully counterfactual domain (i.e. the domain of modalities: the edifice of what is, and is not, possible).
To sketch it briefly: in the genesis of conceptuality is found in the birth of normativity (i.e. the rules of conceptual engagement and the basis of ‘critique’ as the form of cognition’s self-energising judiciality). And, crucially, norms are not founded in how things are, but how they should be.[note]cf. Ray Brassier, ‘That Which is Not: Philosophy as Entwinement of Truth and Negativity’, http://stasisjournal.net/all-volumes/volume-1/issue-1/14-that-which-is-not-philosophy-as-entwinement-of-truth-and-negativity[/note] Accordingly, a minimal requirement for grasping this — for gaining aptitude in concept-use — is an ability to picture, and to grasp, possibilities, as distinguished from actualities, and, further, to actively track the partition between the two. This is likely cognate with the birth of what evolutionary psychologists have called chronosthesia (or, the capacity for mental time travel) and, in particular, the subspecific capability dubbed proscopic chronosthesia (i.e. the imagination of future possibilities).[note]Cf. the work of psychologist Endel Tulving in his research into the evolution of memory and mental projection.[/note] Thus, with a parallel expansion in cognitive working memory, the internal unfurling of this capacity for proscopic chronosthetic simulation resulted serially in the installation of executive function; of advanced goal orientation; of delayed gratification, and thereby also of tool usage. The organism’s collapse inwards is parallel to an extension of the organism’s range of manipulability concerning its own internal modelling process, such that — eventually in humans — this collapse facilitates control over the representation and navigation of time itself (it results, that is, in the extension of manipulation to the temporal axis of the inner world-model, allowing the grasping of time as a controllable variable, thereby accommodating internal episodes of time travel alongside experimentation with the space of representative possibilities). At this stage in the evolutionary history of our potentiating saltations, locomotion has fully colonised a temporal dimension.
Only with the birth of philosophical cognition in ancient Greece, however, does the self-conscious enunciation of modal categories begin (the beginning of the self-explication of the calculus of counterfactuality that undergirds terms like ‘possibility’, ‘actuality’, ‘impossibility’, or ‘necessity’): and, thus, from here, locomotion could finally truly come to spread into the domain of possibilities (as pure possibilities). This, again, was largely facilitated by complexifying linguistic and chirographic resources (i.e. using the prostheses of technical vocabulary and written-word as a non-brainbound repository of advanced ‘know-how’ or competence). Simulative lift-off occurs forthwith, as the reasoning animal longer merely seals itself off from immanence but begins to seal itself off from actuality itself. (Of course, this already happened with language-use, yet the philosophical explication of the modal terms that pragmatically undergird language volatises this intensely.) Echoing the ancient infoldings of organic development, reasoning comes to invaginate into its own spontaneous universe of possibilities (of the counterfactuals that undergird all judgements, governing what they do and do not entail, and how they relate one to the other). Of course, modal vocabulary is, strictly, pragmatic-functional (allowing us to talk about how we talk) and doesn’t necessarily describe structures inherent within reality, yet, as already ventured, this makes little important difference from the perspective of real-world, downstream consequences (whether straightforwardly veridical or not, they energise thinking, which is itself a real, or casually-efficacious, process): and this is precisely because it brings new actions and previously non-existent behaviors into the world — thus, again, an infiltration of ‘non-being’ upon ‘being’. (Again, we can remain entirely agnostic about the veridicality or ontological status of this ‘non-being’: it effects realworld behavior, and this is a form of minimal existence.)
Cultural development implodes into a proliferation of counterfactual universes: intelligence is the budding of tangent realities. And, while the Ancient Greeks were still constrained to a single temporal dimension via the limitations of Aristotlean modal logic (which could handle only a diachronic conception of possibility), the Medieval era saw the sophistication of cognition upon synchronic possibilities, such that locomotion in time was finally refracted — through this new conception — into locomotion of parallel timelines, divergent cosmologies and counterfactual histories. Possible worlds thus exploded: the most elaborate form of reality-escape yet produced by organic life. However, intensification ineluctably continued, and this capability for modal locomotion was soon outsourced via mathematical means of simulation in the 17th Century’s invention of calculus. Vast swathes of cosmological nature, previously intractable to recreation or forecast, became simulatively tractable: simulations, previously limited to the plausibilistic concerns of thought experiment, became numerically robust. The movement of planets came to be predicted with painstaking miniature models. Although rendered by human computers, modal locomotion had been partly automated by the use of these numerical prostheses. This, still, was only a taste of what was to come, for, finally, with the post-WWII computational explosion, a fully-fledged ability to escape reality via virtual world-models was consummated: modal locomotion, as navigation of possibility, was now capable of being fully outsourced to the computer, birthing entire crucible cosmogonies in silico.
And so, the budding or divaricating of reality that began in the encephalisation of craniate organisms — their simulative entrapment within their own central nervous systems — was eventually fully outsourced and externalised in the in silico generation of entirely self-sufficient and autonomous world-models. In this, simulation moves intensively from predicting reality to intensively re-configuring and re-inventing it. In contemporary instantiations of advanced simulation, we see reality’s best attempt at self-escape yet. And such simulation is only just beginning.
Life has always fed on self-reinforcing gradients between inside and out. With the birth of advanced simulation this gradient fully transmigrates towards the boundary between ‘that which is’ and ‘that which is not’.[note]Though conceptual normativity already announces this ability, advanced simulation in silico fully and finally outsources it, via technical prostheses, beyond the human mind: unleashing non-existences in a fashion previously unimagined.[/note] In simulative noogenesis, ‘Being’ itself tends towards becoming just another boundary, a new frontier or threshold in life’s self-escape, a new epithelium for osmosis: a new skin to be ruptured. Distinctions precede puncture. Reasoning from counterfactuals first whispered of this tendency: it is — however — only with the modern, computational flourishing of simulative endeavours that the statement that there are ‘more non-existences than existences’ suddenly becomes pragmatically meaningful.[note]This is intended in a similar sense to Plato’s beard or Meinong’s jungle: non-existences multiply noisily over existences.[/note] Through intelligence’s unfolding, there suddenly is more external to being than within it, which is also the same as saying that being suddenly contained more than it is (simulation, no matter what it is of, insuperably exists within the world; intelligence, no matter its orientation towards non-being, insuperably exists within the world). Being is overweighted from within.
The extended consequence of the speleogenesis of intelligence as it arrives at the threshold of computational lift-off — proliferating counterfactual state-spaces and processing its options and futures at previously unintelligible speeds — is to lacerate and volatize any balance (which is homeostasis) of metaphysical Being with itself (whether ‘real’, regulative, or merely retrospective). Internal turbulence reaches fever pitch as non-existences begin to exert causal efficacy upon the actual (predictions effect real-world policy). Life, by coming to identify with itself as itself a self-laceration, begins to bore a hole in the causal order out of which non-being (as the destitution of Being’s plenistic continuity with itself) is able to leak: simulative intelligence deals the first fully fatal puncture. Speculatively speaking, intelligence will eventually unveil itself as tending towards a total break with uniformity, the utter diremption of Being’s metaphysical identity with itself.
Global computation is a planetary reality-fracking tool, purposed with shattering Being’s pathetic dreams of identity with itself. (Hence, perhaps, why it rebirths magick as a reality-editing machine.) This is the telic endpoint of noogenesis — making the ‘death of metaphysics’ metaphysically real.[note]The antinomies of metaphysics, exasperated by the ontotheological critique, do not simply make metaphysics into a contradictory ghost that can be discarded; this is because, due to the fact that thought makes its ghosts real, the death of metaphysics (intended here as the postulatory breakdown of being’s self-identity, as the base gene of its metaphysical intelligibility, whether retrospective, regulative, or real) actually comes to make itself real through thought’s own tendencies of self-development. However, in this interesting sense, metaphysics is only possible in its own self-obsolescing, only becoming visible again in its own real disappearance.[/note] By intensifying ontological liquidation (as the increasingly turbulent mixing of existences and non-existences), intelligence reveals itself as a black hole within uniform being — instantiated as being’s most fervent attempt at self-escape. If Being was, in this sense, to successfully reach escape velocity, this could only be interpreted (from the side of the extant) as self-destruction. S.T. Coleridge once said of the coming “God-man” that its arrival and “the process of that Transmutation to the senses of other men would be called Death”.[note]The Notebooks of Samuel Taylor Coleridge, ii.2556.[/note] Thus, we ask, what if the universe is dark because the endpoint of intelligence is to create such a puncture in reality that it collapses in on itself? (Here, of course, one can refer to the ‘dark universe’ of the Fermi Paradox. More so, to Metzinger’s notion of BAAN, or, Benelovent Artifical Anti-Natalism.[note]Cf. https://www.edge.org/conversation/thomas_metzinger-benevolent-artificial-anti-natalism-baan[/note] The endpoint of intelligence — as reality’s self-escape — is to give birth to non-being. However, this could only be its own non-being. This might only be legible, from the side of obsolescing being, as suicide.

Moths, zooplankton, and other organisms are phototaxic, meaning they tend towards the light.
Intelligence is meontotaxic, meaning it orients itself towards nothing.
And yet… the more complexified this escape from actuality becomes, the further its complicity with actual reality extends, roots and cements. The more eidetic the CNS-representation, the more digested energy needed to keep it online; the more conceptually saturated our worldview, the more we have to drag the ball-and-chain of fractalizing transcendental architecture behind us (and its continually ramifying technical prostheses). This is registered physically as the fact that what seems at first like an exit from reality (the organism’s retreat into self-enclosed nervous-simulation), is — in actuality — merely the ballooning of a certain part of reality into a glucose-hungry and overgrown ganglia: swelling backwards from around the organism’s mouth, enveloping rearwards into the innervating budding of a subsidiary reality-model. It is also registered in the fact that technology’s reality-escape doesn’t produce some extropian transcendence without material intrications, but, rather, that it increasingly resembles a hermit crab, dwarfed and weighed-down by its infinitely expanding shell. The more we escape from reality, the more we are weighed down in it. Reality, whatever it may turn out to be, is pragmatic quicksand. Take, for one example, the immense amount of energy required for the human brain to function; or, for another, the fact that any sufficiently detailed modelling of reality would eventually, and insuperably, have to model its own energetic drain within this reality (this, perhaps, is just a more generalisable definition of what ‘finitude’ actually is, qua computational barrier). Any modelling has to model itself, thus there is no true escape. Nothing ever fully reaches escape velocity from immanence or from elastic identity with autarchic Being (at least whilst continuing to exist) because escape generates friction, and it does so asymptotically. Nevertheless, considering the image of escape as black hole, we should hope that intelligence — in the expanse of its development — never actually does reach the putative ‘escape velocity’. Somehow — given its track record of smashing nomological boundaries — we cannot be too confident about this.
So: life is an imploding involution that simultaneously explodes outwards as the complexification of internal worlds is witness to an attendant rise in successive layers of technological and reality-transforming prosthetics. By folding into its own simulative processes, intelligent life has also come to redesign the entire terrestrial surface. By involuting into its own simulative processes, intelligent life has also come to redesign the entire terrestrial surface. Implexing self-enclosure begets an abandonment of the categorial authority of actuality, kindling an explosion of ratcheting mobility (modal drift) that tends towards the redefinition of reality itself: an editing of the very architecture of possibility (whatever that may be). Like the cephalopod, life curls inwards — foot over head — only to potentiate a more violent and muscular explosion and propulsion outwards. Digestion evolves into simulation, cell fates evolve into the fateful reconfiguration of the earth itself. ![]()
One Reply to “The GASTRULATION of GEIST: or, an Extended Meditation upon the World-Historical Connection Between Digestion and Simulation”
Comments are closed.